欢迎光临~北京凯森莱科技有限公司
语言选择:
 ∷
∷

∷
 ∷
∷
Middle East respiratory syndrome coronavirus (MERS-CoV) is the causative agent of a severe respiratory disease associated with more than 2468 human infections and over 851 deaths in 27 countries since 2012. There are no approved treatments for MERS-CoV infection although a combination of lopinavir, ritonavir and interferon beta (LPV/RTV-IFNb) is currently being evaluated in humans in the Kingdom of Saudi Arabia. Here, we show that remdesivir (RDV) and IFNb have superior antiviral activity to LPV and RTV in vitro. In mice, both prophylactic and therapeutic RDV improve pulmonary function and reduce lung viral loads and severe lung pathology. In contrast, prophylactic LPV/RTV-IFNb slightly reduces viral loads without impacting other disease parameters. Therapeutic LPV/RTV-IFNb improves pulmonary function but does not reduce virus replication or severe lung pathology. Thus, we provide in vivo evidence of the potential for RDV to treat MERS-CoV infections.
The coronavirus (CoV) family has a propensity for emergence into new hosts often causing novel severe disease. In 2012, Middle East respiratory syndrome coronavirus (MERS-CoV), was discovered as the causative agent of a severe respiratory syndrome in the Kingdom of Saudi Arabia (KSA), has since caused at least 2468 cases and 851 deaths globally1. MERS-CoV is endemic in camels, the zoonotic reservoir host, with evidence of infection going back at least 30 years2. Camels in the Middle East and perhaps in East Africa continue to seed human infections which may require hospitalization especially in aged individuals with preexisting comorbidities1,3,4. Similar to severe acute respiratory syndrome CoV (SARS-CoV), MERS-CoV has spread to over 27 countries via air travel of infected people5. In 2014, a single imported case caused an outbreak of 186 cases in South Korea, while a more recent case imported from the Middle East was contained as a result of rapid implementation of public health measures6. MERS-CoV continues to cause human infections globally and thus is listed as a priority pathogen with pandemic potential by World Health Organization (WHO) and the Coalition for Epidemic Preparedness Innovations (CEPI). Presently, there are no approved treatments for MERS-CoV or any other human CoV.
Emerging viral diseases typically have very few if any effective treatment options. As such, treatments designed and approved for other diseases are administered to patients with emerging viral syndromes empirically based on limited clinical or laboratory data. Multiple U.S. Food and Drug Administration (FDA) approved therapies have been evaluated for antiviral activity against MERS-CoV in vitro including lopinavir (LPV), ritonavir (RTV), and interferon beta (IFNb). LPV is a human immunodeficiency virus 1 (HIV-1) protease inhibitor that is usually combined with RTV to increase LPV half-life through the inhibition of cytochrome P4507. Although the antiviral activity of LPV against MERS-CoV has been reported in Vero cells (concentration causing a 50% reduction in replication (EC50) = 8 µM), other studies report complete inactivity similar to that of RTV8,9. In contrast, studies evaluating the antiviral activity of type I and type II interferons have reported IFNb as the most potent interferon (EC50 1.37–17 IU/mL) in reducing MERS-CoV replication in vitro8,10. The only in vivo study assessing the therapeutic efficacy of LPV/RTV or IFNb against MERS-CoV published thus far was performed in common marmosets where modest improvements in clinical outcomes were noted11. In human MERS-CoV patients, two published case reports describe conflicting results on the use of a combination of LPV/RTV, pegylated interferon, and ribavirin with one of two patients surviving12,13. To this end, a randomized control trial (MIRACLE Trial) aimed at conclusively determining if LPV/RTV-IFNb improves clinical outcomes in MERS-CoV patients was initiated in 2016 and has thus far enrolled 76 patients in KSA14,15.
Remdesivir (RDV, GS-5734) is a broad-spectrum antiviral nucleotide prodrug with potent in vitro antiviral activity against a diverse panel of RNA viruses such as Ebola virus (EBOV), Marburg, MERS-CoV, SARS-CoV, respiratory syncytial virus (RSV), Nipah virus (NiV), and Hendra virus16,17,18. The mechanism of RDV’s anti-MERS-CoV activity is likely through premature termination of viral RNA transcription as shown in biochemical assays using recombinant EBOV, NiV, and RSV polymerases18,19,20. In primary human lung epithelial cell cultures, RDV is potently antiviral against circulating contemporary human CoVs, SARS-CoV (EC50 = 0.07 µM), MERS-CoV (EC50 = 0.07 µM), and related zoonotic bat CoVs17,21. We recently reported that therapeutic RDV improves disease outcomes and reduces viral loads in SARS-CoV-infected mice17. Since similar studies had not been performed with MERS-CoV, we generated a transgenic mouse with a humanized MERS-CoV receptor (dipeptidyl peptidase 4, hDPP4) and deleted for carboxylesterase 1c (Ces1c) to improve the pharmacokinetics of nucleotide prodrugs such that it better approximates the drug exposure profile in humans22. Here, we show that RDV provides superior antiviral activity against MERS-CoV in vitro and in vivo as compared with LPV/RTV-IFNb. In addition, RDV was the only therapeutic treatment to significantly reduce pulmonary pathology. Thus, we provide in vivo evidence of the potential for RDV to treat MERS-CoV infections.
We utilized a recombinant MERS-CoV engineered to express a reporter nanoluciferase (MERS-nLUC) for our in vitro antiviral activity assays. To ensure that our reporter virus behaved similarly to WT MERS-CoV, we first demonstrated that MERS-nLUC and wild-type (WT) MERS-CoV EMC 2012 strain replicate to similar levels in the absence of drug treatment and are similarly susceptible to the antiviral activity of RDV (WT EC50 = 0.12 µM; MERS-nLUC EC50 = 0.09 µM) in the human lung epithelial cell line, Calu-3 (Supplementary Fig. 1). These data are in agreement with those for WT MERS Jordan strain (EC50 = 0.3 µM) reported by Warren et al.18. Thus, future reporter virus data should be representative of WT MERS-CoV.
We then performed parallel antiviral assays in Calu-3 cells with MERS-nLUC comparing LPV, RTV, IFNb, and RDV (Fig. 1; Supplementary Fig. 2)17. Similar to our previous reports, RDV showed potent inhibition of MERS-CoV replication with a EC50 of 0.09 µM, no observable cytotoxicity up to 10 μM and a selectivity index (SI = EC50/CC50) > 10017. (Fig. 1a). In contrast, the respective EC50 values generated for LPV and RTV were 11.6 and 24.9 µM with CC50 values >50 µM (Fig. 1b, c). Thus, the SI for LPV and RTV was > 4.3 and > 2, respectively. Combination LPV and RTV (LPV:RTV, 4.6:1 molar ratio) is currently under evaluation in the MIRACLE trial14. The antiviral activity of LPV/RTV (EC50 = 8.5 µM) was similar to LPV alone (EC50 = 11.6 µM, P = 0.43, Wilcoxon matched-pairs signed rank test), suggesting the effect was largely driven by LPV (Fig. 1d). We found potent inhibition of MERS-CoV with IFNb (EC50 = 175 international units (IU)/mL) (Fig. 1e), CC50 values >2800 IU/mL and an SI > 16. Together, these data demonstrate that RDV and IFNb have superior in vitro antiviral activity compared with LPV and RTV, and that RTV does not significantly enhance the antiviral activity of LPV in vitro.
Fig. 1: RDV and IFNb have superior antiviral activity to LPV and RTV.
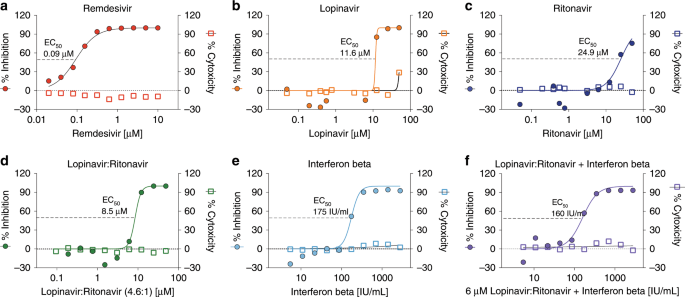
Graphs depict mean % inhibition of MERS-CoV replication (left Y-axis) and % cytotoxicity (right Y-axis) of antivirals. Calu-3 cells were infected in sextuplicate with MERS-CoV nanoluciferase (nLUC) at a multiplicity of infection (MOI) of 0.08 in the presence of a dose response of drug for 48 h, after which replication was measured through quantitation of MERS-CoV–expressed nLUC. Cytotoxicity was measured in similarly treated but uninfected cultures via Cell-Titer-Glo assay. Representative data are shown from four independent experiments.
Cell culture medium and human plasma have different concentrations and types of proteins, which directly affects the levels of biologically available free drug unbound to protein complicating the comparison of drug levels the respective systems23. To address this issue, we utilized comparative equilibrium dialysis (CED) to determine the differences in free unbound drug between human plasma and cell culture medium revealing that the maximal plasma concentration (Cmax) in human plasma (15 µM) had the same amount of free unbound LPV as 5 µM LPV in 10% FBS containing cell culture medium. Thus, we combined LPV (5 µM) and RTV (1.09 µM) at a fixed molar ratio of 4.6:1 in combination with increasing concentrations of IFNb. The antiviral activity of the LPV/RTV-IFNb combination (EC50 = 160 IU/mL) was indistinguishable from that of IFNb alone (EC50 = 175 IU/mL) (P = 0.62, Wilcoxon matched-pairs signed rank test) (Fig. 1f). These data suggest that the observed in vitro antiviral activity of the LPV/RTV-IFNb combination on MERS-CoV is dominated by IFNb when LPV/RTV is used at clinically relevant concentrations.
Unlike humans, mice have high levels of a serum esterase (carboxylesterase 1c, Ces1c) that drastically reduces the stability of RDV in mice requiring efficacy studies be performed in Ces1c−/− mice to better approximate the pharmacokinetics and drug exposure profile in humans17. MERS-CoV infection of standard laboratory mice is prevented due to differences in human and mouse dipeptidyl peptidase 4 (DPP4), the entry receptor for MERS-CoV. To enable testing of RDV in mice, we bred Ces1c−/− mice with mice harboring a modified DPP4 humanized via CRISPR/Cas9 at residues 288 and 330 (hDPP4). The resultant Ces1c−/−hDPP4 mice had indistinguishable virus replication and pathogenesis from hDPP4mice when infected with MERS-CoV (Supplementary Fig. 3)22,24.
Using the new Ces1c−/− hDPP4 mouse model, we sought to determine if prophylactic RDV could ameliorate MERS-CoV disease. As shown in Fig. 2a, prophylactic RDV (25 mg/kg, BID) administered 1 day prior to infection significantly diminished MERS-CoV-induced weight loss in mice infected with 5E + 04 (P < 0.0001, two-way ANOVA with Tukey’s multiple comparison test) or 5E + 05 plaque-forming units (pfu) (P < 0.0001, two-way ANOVA with Tukey’s multiple comparison test) as compared with similarly infected vehicle-treated animals. Moreover, RDV administered prior to infection also prevented mortality (P = 0.0037, Mantel–Cox test) in those administered a lethal dose (i.e., 5E + 05 pfu) (Fig. 2b). In contrast to vehicle-treated animals, lung hemorrhage was significantly reduced (P < 0.0001, two-way ANOVA with Tukey’s multiple comparison test) with RDV prophylaxis (Fig. 2c)22,25,26. Importantly, RDV prophylaxis significantly reduced virus lung titers > 3 logs on both 4 (5E + 04 P = 0.0240, 5E + 04 P = 0.0001, two-way ANOVA with Sidek’s multiple comparison test) and 6 days post infection (dpi) (5E + 05 P = 0.0001, two-way ANOVA with Sidek’s multiple comparison test) (Fig. 2d), which was corroborated by viral antigen labeling in lung tissue sections (Fig. 2e).
Fig. 2: Prophylactic RDV reduces MERS-CoV replication and disease.
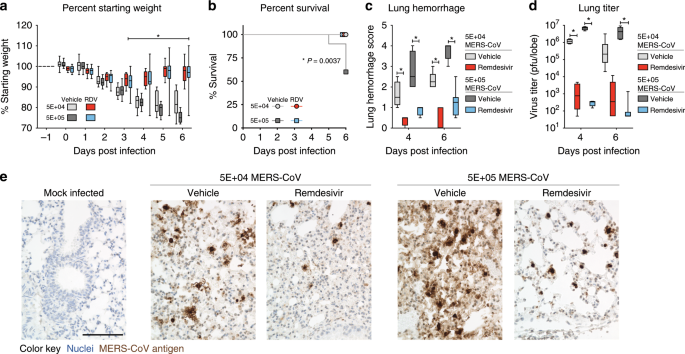
a Percent starting weight of 9–12-week-old male and female Ces1c−/− hDPP4 mice prophylactically administered subcutaneous vehicle or remdesivir (RDV, 25 mg/kg) BID the day prior to infection with either 5E + 04 (vehicle n = 14, RDV n = 14) or 5E + 05 (vehicle n = 14, RDV n = 15) plaque-forming units (pfu) MERS M35C4. Asterisks indicate statistically significant differences (P < 0.05) as determined by two-way ANOVA and Tukey’s multiple comparison test. b Percent survival of each cohort and survival analysis by Mantel–Cox test (P < 0.05, N per group noted in a). c Lung hemorrhage scored on a scale of 0–4, where 0 is a normal pink healthy lung and 4 is a completely dark red lung. On 4 dpi, N = 4/group, and on 6 dpi the remaining animals are plotted. Asterisks indicate statistically significant differences (P < 0.05) as determined by two-way ANOVA and Tukey’s multiple comparison test. d MERS-CoV lung titer on 4 (N = 4) and 6 dpi (all remaining animals). Asterisks indicate statistically significant differences (P < 0.05) as determined by two-way ANOVA and Sidek’s multiple comparison test. For a, c, d, the boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range. e Hematoxylin (nuclei, blue) and immunostaining for MERS-CoV antigen (brown) in lung tissue sections from 4 dpi. All photos were taken with the same magnification. The black bar indicates 100 µM scale. Images from representative mice for each group are shown.
The American Thoracic Society (ATS) issued a consensus document with defined parameters and tools to more accurately translate small animal models of acute lung injury (ALI) to the human condition26. Using the ATS Lung Injury Scoring System designed to quantitate histopathological features of ALI (Fig. 3a), we blindly scored fields of hematoxylin and eosin stained lung tissue sections from the mice in Fig. 2 for the following features: neutrophils in the alveolar and interstitial space, hyaline membranes, proteinaceous debris filling the air spaces, and alveolar septal thickening26. As compared with control mice, the ATS lung injury scores were significantly reduced in RDV-treated mice (P = 0.0079, Mann–Whitney test) infected with 5E + 04 pfu MERS-CoV and approached statistical significance (P = 0.0536, Mann–Whitney test) in those infected with the higher virus dose (5E + 05 pfu) (Fig. 3a). In Fig. 3, we also show examples of the quantitated pathological features. In normal healthy mock-infected control mice (Fig. 3b), the alveolar air spaces are free of debris and inflammatory cells, the walls (i.e., septae) of the alveolar sac are thin which facilitates efficient gas exchange and rare neutrophils in circulation are seen in the capillaries within alveolar septae, but not in the air spaces. In contrast, vehicle-treated MERS-CoV-infected animals in both viral dosage groups (Fig. 3c) had multiple histologic features of ALI including notable immune cell infiltration into the alveolar septae and resultant septal wall thickening, scattered degenerating and dying cells, proteinaceous debris in the air spaces resulting from capillary leakage some of which is organized into hyaline membranes and neutrophils in alveolar septae as well as in the air spaces. Interestingly, the early hyaline membranes (i.e., thin pink material lining alveoli) were noted in vehicle-treated animals infected with the lower dose of MERS-CoV (Fig. 3c, left), while more developed hyaline membranes (i.e., thick pink material lining alveoli) were observed in vehicle-treated animals infected with the higher dose of virus (Fig. 3c, right). Importantly, the air spaces of RDV-treated animals infected with the lower dose of MERS-CoV (Fig. 3d, left) predominantly lacked cellular debris, immune cells, and hyaline membranes although alveolar septal thickening due to increased immune cells was observed. RDV-treated animals infected with the higher dose of MERS-CoV (Fig. 3d, right) similarly lacked debris and inflammatory infiltrates in the air spaces, but exhibited increased alveolar septal thickening. Thus, our blind pathological evaluation of lung tissue sections using the ATS Lung Injury Scoring System demonstrates that prophylactic RDV diminished the pathological features of ALI in MERS-CoV-infected mice26.
Fig. 3: Remdesivir prophylaxis reduces features of acute lung injury.
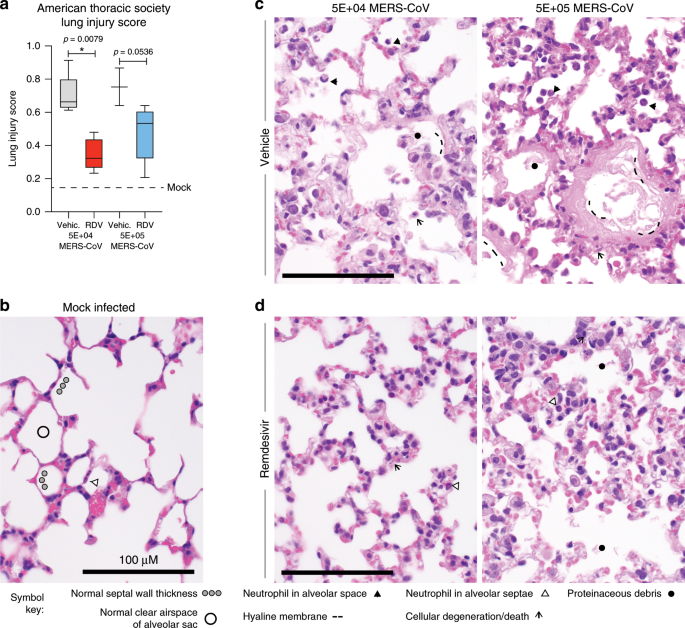
The histological features of acute lung injury were blindly scored using the American Thoracic Society Lung Injury Scoring system creating an aggregate score for the following phenotypes: neutrophils in the alveolar and interstitial space, hyaline membranes, proteinaceous debris filling the air spaces, and alveolar septal thickening. Three randomly chosen high power (×60) fields of diseased lung were assessed per mouse. Representative images are shown for mock infected as well as those administered prophylactic vehicle or RDV and infected with either 5E + 04 or 5E + 05 pfu mouse adapted MERS-CoV. The numbers of mice scored per group: Vehicle 5E + 04 pfu MERS-CoV N = 5, Vehicle 5E + 05 pfu MERS-CoV N = 3, RDV 5E + 04 pfu MERS-CoV N = 5, RDV 5E + 05 pfu MERS-CoV N = 5. Symbols identifying example features of disease are indicated in the figure. All images were taken at the same magnification. The black bar indicates 100 µm scale. For the graph, the boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range. Statistical significance was determined by Mann–Whitney test.
Prophylactic studies provide a best case scenario to evaluate in vivo antiviral activity since time is given for the metabolization and accumulation of antiviral agents within cells targeted by virus prior to infection. To determine if prophylactic LPV/RTV-IFNb improved outcomes following MERS-CoV infection, we first confirmed that subcutaneous human equivalent doses (H.E.D.) of IFNb exerted a biological effect in mice. Administration of a 1 × , 2.5x, or 25x H.E.D. of mouse IFNb rapidly induced dose-dependent expression of interferon stimulated gene (ISG), Mx1, in peripheral blood mononuclear cells (PBMCs) in Ces1c−/− mice (Supplementary Fig. 4a). Importantly, the kinetics of Mx1 gene expression in target organ of MERS-CoV, the lung, was similar in both Ces1c−/− and Ces1c−/− hDPP4mice (Supplementary Fig. 4b). Similarly, a single dose of IFNb significantly induced sustained expression (P < 0.05, two-way ANOVA with Sidek’s multiple comparison test) of interferon gamma-induced protein 10 (IP-10, CXCL-10) in the serum of both strains of mice (Supplementary Fig. 4c). Thus, we observed an expected ISG response in the blood and lung tissue of both Ces1−/− and Ces1−/−hDPP4 mice following IFNb treatment.
We then evaluated if LPV/RTV-IFNb prophylaxis could improve outcomes in Ces1−/− hDPP4 mice infected with 5E + 04 pfu MERS-CoV. We compared vehicle (oral: propylene glycol and ethanol, subcutaneous: PBS) to three different treatment scenarios, including LPV/RTV-IFNb high (25x H.E.D. IFNb), LPV/RTV-IFNb low (1x H.E.D. IFNb), or IFNb-high alone (25x H.E.D. IFNb) (Fig. 4). Prophylactic RDV and vehicle were included as controls. Since ISG expression peaks 2–4 h after IFNb administration (Supplementary Fig. 4), we initiated IFNb dosing 2 h prior to MERS-CoV infection to maximize the potential antiviral effect. Similar to our previous studies, RDV (25 mg/kg, BID) or vehicle were administered subcutaneously every 12 h to obtain exposures in mice similar to that observed in humans17. The dosing levels and frequencies of LPV/RTV (oral once daily) and IFNb (subcutaneously every other day) were chosen to mirror those in the MIRACLE trial27. Unlike RDV-treated mice (Fig. 4a), vehicle, LPV/RTV-IFNb, or IFNb alone did not prevent weight loss (Fig. 4b). In fact, animals administered IFNb alone lost significantly more weight than vehicle (P = 0.01), LPV/RTV-IFNb high (P = 0.0001) and low (P = 0.006, all two-way ANOVA with Tukey’s multiple comparison test) groups (Fig. 4b). On 6 dpi, RDV treatment reduced virus lung titers the most (>3 log reduction, vehicle median = 1.4E + 05 pfu/lobe, RDV median = 50 pfu/lobe) (Fig. 4a), while those in LPV/RTV-IFNb low treated were modestly reduced (~4-fold, median = 5.5E + 03 pfu/lobe) as compared with its vehicle (median = 2.3E + 04 pfu/lobe) (P = 0.04, two-way ANOVA with Sidek’s multiple comparison test) (Fig. 4b). Upon measuring pulmonary function by whole-body plethysmography (WBP), we found only RDV prophylaxis improved pulmonary function (Fig. 4a, b) (P = 0.002 to 0.0001, two-way ANOVA with Sidek’s multiple comparison test). Interestingly, animals receiving IFNb-alone had significantly worse pulmonary function than companion groups at later times post infection (4–5 dpi, P = < 0.05, two-way ANOVA with Tukey’s multiple comparison test). To determine if initiation time or dose level would affect outcomes in the IFNb only group, we performed a similar study but administered IFNb at a lower dose (i.e., 1x H.E.D.) 24 h prior to infection (Supplementary Fig. 5). Alteration of IFNb dose or time of administration did not improve outcomes from those in Fig. 4, and the modest improvement in virus titer seen with LPV/RTV-IFNb noted above was not observed. Taken together, prophylactic LPV/RTV-IFNb caused modest reductions in lung viral load in one of two studies, but had minimal impact on other disease metrics while IFNb alone did not impact virus replication and exacerbated disease.
Fig. 4: Prophylactic LPV/RTV + IFNb does not improve disease outcomes.
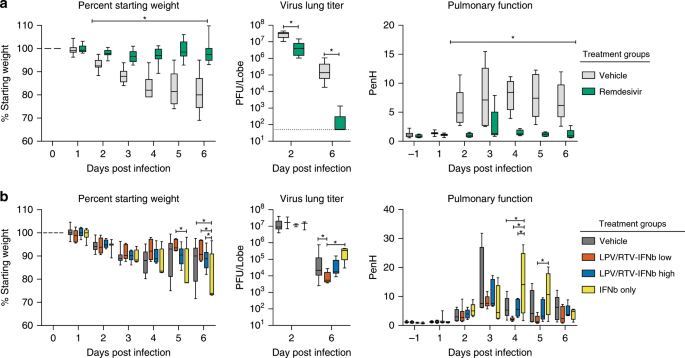
a Percent starting weight (Left) of 12–14-week-old female Ces1c−/− hDPP4 mice infected with 5E + 04 pfu MERS M35C4 and treated BID with either vehicle (n = 9) or remdesivir (RDV, 25 mg/kg, n = 9) subcutaneously beginning −1 dpi. Asterisks indicate statistically significant differences (P < 0.05) as determined by two-way ANOVA and Tukey’s multiple comparison test. (Middle) MERS-CoV lung titer on 2 (N = 3) and 6 dpi (all remaining animals). Asterisks indicate statistically significant differences (P < 0.05) as determined by Mann–Whitney test. (Right) WBP was used to assess pulmonary function in mice. PenH is a surrogate measure of airway resistance or bronchoconstriction. Asterisks indicate statistical differences by two-way ANOVA with Sidek’s multiple comparison test. b Percent starting weight (left), virus lung titer (middle), and pulmonary function metric PenH (right) of cohorts of mice similar in age and sex and infected similarly with MERS-CoV as in b but treated with vehicle (n = 9), LPV/RTV + IFNb low (1× human equivalent) (n = 9), LPV/RTV + IFNb high (25× human equivalent) (n = 9), or IFNb high only (n = 9). Oral vehicle or lopinavir/ritonavir (160/40 mg/kg) were administered orally once daily beginning the −1 dpi. IFNb treatment was initiated 2 h prior to infection and every other day thereafter. To control for dosing effects, vehicle-treated mice received both LPV/RTV vehicle and subcutaneous PBS to mirror IFNb injections. Likewise, IFNb only group received oral vehicle to mirror that seen in orally dosed groups. Similar statistical tests performed on awere performed on b. For the box and whisker plots, the boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range.
To more stringently assess the therapeutic potential of RDV and LPV/RTV-IFNb and to better model the human scenario where MERS-CoV patients most likely would initiate treatment after infection, we performed a series of therapeutic efficacy studies in mice. We initiated the following treatments in Ces1−/− hDPP4mice infected with 5E + 04 pfu MERS-CoV on 1 dpi: RDV or vehicle, LPV/RTV-IFNb low (1× human equivalent), LPV/RTV-IFNb high (25× human equivalent) or their vehicles. Dose route, amount, and frequency were similar to the prophylactic studies above. Only therapeutic RDV substantially reduced body weight loss (P = 0.019 to < 0.0001, two-way ANOVA with Tukey’s multiple comparison test) (Fig. 5a, b) and lung hemorrhage on 6 dpi (Fig. 5c) (P < 0.0001, one-way ANOVA with Kruskal–Wallis test). Similarly, only RDV treatment significantly reduced virus lung titers on 6 dpi (vehicle median 7.8E + 05 pfu/lobe, RDV median 125 pfu/lobe, P = 0.0001, one-way ANOVA with Kruskal–Wallis test) (Fig. 5d), which we corroborated with viral antigen labeling in lung tissue sections (Fig. 5e). Similar therapeutic studies (Supplementary Fig. 6) were performed but with a lethal dose of MERS-CoV (5E + 05 pfu) where no treatment improved survival (Supplementary Fig. 6a, d) or lung hemorrhage (Supplementary Fig. 6b, e), but therapeutic RDV significantly reduced lung viral load on 6 dpi (Supplementary Fig. S6c) (P = 0.03, Mann–Whitney test). Thus, therapeutic LPV/RTV-IFNb failed to improve weight loss, lung hemorrhage, and virus lung titer after 5E + 04 pfu MERS-CoV and did not improve survival following a lethal dose of MERS-CoV (Supplementary Fig. 6d–f). In contrast, therapeutic RDV diminished weight loss, lung hemorrhage, and virus replication during an ongoing MERS-CoV infection, but the degree of clinical benefit is dependent on viral dose and time of treatment initiation.
Fig. 5: Therapeutic RDV reduces replication and pathology.
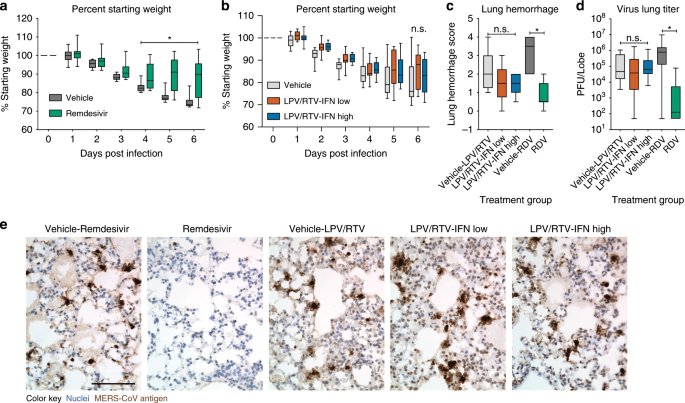
Percent starting weight of 10–12-week-old female Ces1c−/− hDPP4 mice infected with 5E + 04 pfu MERS M35C4 and treated with a subcutaneous vehicle for RDV (N = 13) or remdesivir (RDV, 25 mg/kg, N = 14) BID beginning 1 dpi or b vehicle for LPV/RTV-IFNb (N = 15), LPV/RTV-IFNb low (N = 16) or LPV/RTV-IFNb high (N = 16) beginning 1 dpi. Oral vehicle or lopinavir/ritonavir (160/40 mg/kg) was administered orally once daily. IFNb low (1x human equivalent dose of 1.6 MIU/kg) and high (25x human equivalent dose of 40 MIU/kg) or PBS vehicle were administered via subcutaneous injection every other day. Asterisks indicate statistical differences by two-way ANOVA with Tukey’s multiple comparison test. c Lung hemorrhage 6 dpi for all animals in a, b scored on a scale of 0–4, where 0 is a normal pink healthy lung and 4 is a diffusely discolored dark red lung. d MERS-CoV lung titer 6 dpi in mice as described in a, b. Asterisks indicate statistical significance (N group described in a and b, P < 0.05) by one-way ANOVA with Kruskal–Wallis test for (c, d). Data for a–d are compiled from two independent experiments. For the box and whisker plots, the boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range. e Representative photomicrographs of MERS-CoV antigen (brown) and hematoxylin stained nuclei (blue) in mouse lung tissue sections from 6 dpi. The black bar is 100 µM.
In the therapeutic antiviral efficacy studies described in Fig. 5, we used WBP to assess pulmonary function (Fig. 6). In contrast to vehicle-treated animals, RDV-treated animals had reduced flow rate at 50% of the expired volume (EF50) (P = 0.01, two-way ANOVA with Sidek’s multiple comparison test), and PenH (P = 0.04, two-way ANOVA with Sidek’s multiple comparison test), a surrogate measure of airway resistance/obstruction (Fig. 6a)28. Similarly, Rpef (the fraction of expiration before peak expiratory flow is reached), an indicator of bronchoconstriction, returned to baseline in RDV-treated animals by 5 dpi (P = 0.002, two-way ANOVA with Sidek’s multiple comparison test) yet remained suppressed in vehicle-treated animals. Unlike the prophylactic study in Fig. 4b, therapeutic LPV + RTV-IFNb low improved pulmonary function as compared with vehicle with significantly reduced EF50 and PenH (P < 0.05, two-way ANOVA with Tukey’s multiple comparison test) and baseline levels of Rpef by 5 dpi (P < 0.05, two-way ANOVA with Tukey’s multiple comparison test) (Fig. 6b). While therapeutic LPV/RTV-IFNb failed to reduce weight loss, lung hemorrhage, and virus titer, this regimen appears to provide improvements in pulmonary function similar to therapeutic RDV.
Fig. 6: Therapeutic RDV and LPV/RTV-IFNb improve pulmonary function.
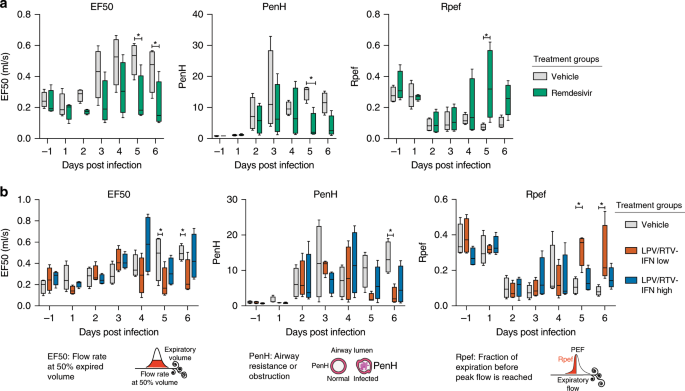
Whole-body plethysmography (WBP) was used to assess pulmonary function in mice. Representative WBP data for one of the two studies depicted in Fig. 5 are shown. All groups were N = 4 per day. EF50 is the flow rate at 50% expired volume. PenH is a surrogate measure of airway resistance. Rpef is the fraction of expiration before peak expiratory flow is reached. Altered EF50, PenH, and Rpef indicate bronchoconstriction or airway obstruction. Asterisks indicate statistical differences by two-way ANOVA with Sidek’s multiple comparison test in a and two-way ANOVA with Tukey’s multiple comparison test in b. The boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range.
We then quantitated the lung pathology in therapeutically treated animals (Fig. 7). In lung tissue sections from the 6 dpi, we blindly evaluated and scored features of ALI using three different and complementary approaches. In Fig. 7a, we show reduced ALI with RDV treatment. Although sporadic apoptotic cells and alveolar septal wall thickening (~2–4-fold over mock) driven by immune cell infiltration were observed in the lungs of mice treated with therapeutic RDV, the air spaces remained predominantly free of cellular debris and inflammatory cells similar to mock-infected animals (Fig. 7a). In contrast, multiple pathological features typical of ALI were noted in both vehicle groups (RDV and LPV/RTV vehicles) as well as both LPV/RTV-IFNb low and high, including altered alveolar architecture due to pneumocyte degeneration and death, numerous inflammatory cells in the septae and in alveolar air spaces, neutrophils in the air spaces, and proteinaceous debris in the air spaces organizing into hyaline membranes (Fig. 7a). Using the ATS ALI scoring tool described in Fig. 3, we found that only RDV therapy significantly (P = 0.005, one-way ANOVA with Kruskal–Wallis test) decreased lung injury scores (Fig. 7b). With a complementary histological tool, we then quantitated features of diffuse alveolar damage (DAD), the pathological hallmark of ALI26. We found that only therapeutic RDV reduced DAD scores (P = 0.04, one-way ANOVA with Kruskal–Wallis test) (Fig. 7c)29. Since degree of cell death appeared to correlate with protective efficacy, we quantitated levels of cleaved caspase-3 in lung tissue sections by antibody labeling. Caspase-3, a widely accepted marker of apoptosis, is a regulatory enzyme whose cleavage and activation drive programmed cell death30. Using the Definiens software suite, we obtained unbiased quantitative data showing that only RDV treatment significantly reduced levels of cleaved caspase-3 antigen (P = 0.0109, one-way ANOVA with Kruskal–Wallis test) (Fig. 7d). Thus, using three complementary and blinded approaches, we obtained similar data showing that only therapeutic RDV reduced histologic features of ALI.
Fig. 7: Therapeutic RDV but not LPV/RTV-IFNb diminishes signs of ALI.
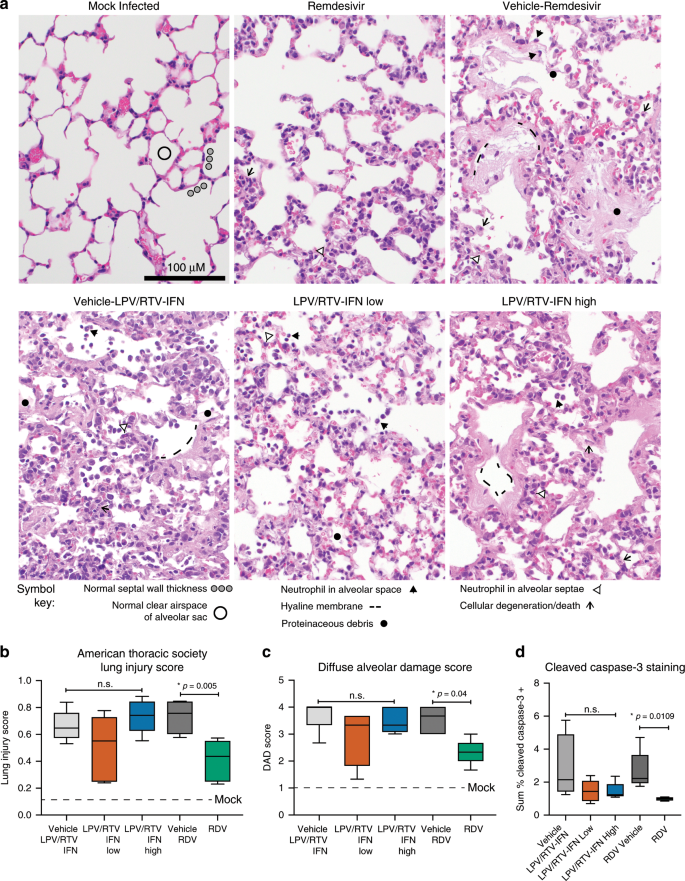
a Representative images of the histological features of acute lung injury 6 dpi comparing a mock-infected mouse to the therapeutic treatment groups described in Figs. 5 and 6. Symbols identifying example features of disease are indicated in the figure. b American Thoracic Society Lung Injury Score derived as described in Fig. 3. The numbers of animals per group quantitated: vehicle RDV N = 7, RDV N= 7, vehicle LPV/RTV-IFNb N = 9, LPV/RTV-IFNb low N= 7, LPV/RTV-IFNb high N = 8. c Diffuse alveolar damage score quantitating the degree of cellular sloughing, necrosis, and breakdown of barrier epithelium and vascular leakage. For both b and c, scores were blindly assessed in three random high power (×60) fields of diseased lung tissue sections. d Quantitation of cleaved caspase-3 antigen staining in lung tissue sections from studies described in Figs. 5–7. Cleaved caspase-3 is a marker of cell death. The numbers of animals per group quantitated for all groups was N = 5/group. For the box and whisker plots, the boxes encompass the 25th to 75th percentile, the line is at the median, while the whiskers represent the range. For b–d, asterisks indicate statistical significance by one-way ANOVA and Kruskal–Wallis multiple comparison test.
Emerging viral diseases have caused significant global pandemics (e.g., HIV, 1918 influenza, smallpox), epidemics (e.g., SARS-CoV), and devastating outbreaks (e.g., EBOV, MERS-CoV). For CoV, metagenomic studies in wild animals have revealed a great diversity of viruses and hosts, and have even identified viruses similar to current and past epidemic strains in bats31,32. Thus, broad-spectrum therapies effective against known epidemic and zoonotic strains likely to seed future emergence, have the potential to diminish epidemic disease today and diminish future outbreaks. Currently, there are no FDA-approved treatments for any human CoV infection. Upon emergence of SARS- and MERS-CoV, patients were administered off-label antivirals (e.g., ribavirin, LPV, RTV) and immunomodulators (e.g., corticosteroids, interferon alpha-2a/2b, IFNb) as single agents or in combination in an attempt to ameliorate severe disease outcomes with very limited success33. Without randomized controlled trials, determining efficacy is difficult due to patient and treatment variability as well as a lack of appropriate matching controls. Although recent meta-analysis and modeling has suggested that interferon treatment does not improve clinical outcomes in MERS-CoV patients4, the MIRACLE trial in the KSA is aimed at conclusively determining if a fixed dose combination of LPV/RTV-IFNb is effective at treating MERS-CoV infections14. Here, we show that RDV provides superior antiviral activity against MERS-CoV in vitro and in vivo as compared with LPV/RTV-IFNb. In addition, RDV was the only treatment to significantly reduce pulmonary pathology. Thus, we provide in vivo evidence of the potential for RDV to treat MERS-CoV infections. Efficacy testing of antiviral regimens in humans like those in the MIRACLE trial are essential to progress antiviral development and prioritize therapies most likely to improve clinical outcomes in MERS-CoV patients.
Differences in protein identity and concentration complicate the translation of in vitro antiviral activity to in vivo therapeutic efficacy. Although the EC50 for LPV against MERS-CoV (EC50 = 11 µM) falls within the maximum (Cmax = 15 µM) and minimum (Cmin = 9.5 µM) levels observed in human plasma, the biologically available fraction unbound to protein in these respective systems should be compared to more accurately translate in vitro activity to potential in vivo efficacy9. For example, the trough levels (Cmin) of total (5.8 µM) and unbound protein free LPV (0.057 µM) in women with HIV differ by 100-fold, but LPV is still highly active and effective since the EC50 (0.010–0.027 µM) falls below the unbound protein free Cmin7,34. Together, these data argue that levels of free biologically active LPV achieved in humans are well below those that exert robust antiviral effects on MERS-CoV replication in cell culture systems. Thus, the micromolar EC50 observed for LPV against MERS-CoV coupled with protein binding and insufficient levels of free LPV in plasma is likely responsible for the modest antiviral effect with LPV/RTV-IFNb prophylaxis and minimal impact on disease with therapeutic administration in our mouse model. In contrast, human equivalent doses of RDV are demonstrably efficacious in mice infected with SARS- and MERS-CoV and in nonhuman primates infected with EBOV thus demonstrating a more suitable PK/PD relationship17.
Interferons are useful in treating multiple viral infections35,36. Since IFNb was shown to be the most potent against MERS-CoV when comparing the antiviral activity of multiple type I and type II interferons in Vero cells, IFNb was selected to use in the MIRACLE trial8,10. Mirroring the MIRACLE trial, we delivered IFNb subcutaneously every other day, which failed to reduce MERS-CoV viral loads and appeared to exacerbate disease in mice. As we sought to understand this result, we found little experimental congruency among reports detailing the prophylactic or therapeutic efficacy of type I interferons in animal models of MERS-CoV11,37,38. Falzarano et al. demonstrated that IFN-alpha-2a coupled with ribavirin initiated 8 h post infection improved outcomes in rhesus macaques and reduced viral copy number in lung tissue, but the treatment had no effect on infectious virus titers in bronchoalveolar lavage fluid37. In mice where hDPP4 is delivered to lung tissue by adenoviral transduction, intranasal IFNb given before or after MERS-CoV infection reduced lung titers although the peak lung titers in this model are approximately two orders of magnitude lower than the current transgenic models, and thus may be more easily treated38,39,40. The utility of the common marmoset as a model of MERS-CoV pathogenesis is controversial with one study detailing severe respiratory disease yet another reporting similarly mild disease among mock and MERS-CoV-infected animals41,42. In marmosets, Chan et al. explored the therapeutic potential of LPV/RTV or IFNb, but the small numbers of animals used per group, lack of time-matched viral load samples, and unexpected early mortality in the LPV group made the resultant data difficult to interpret11. Nevertheless, the studies noted above demonstrate that type I interferon can exert an antiviral effect on MERS-CoV in vivo when given subcutaneously (IFN alpha, rhesus macaque) and intranasally (IFNb, adenovirus hDPP4 model)37,38,39. Our inability to reduce MERS-CoV titer or improve outcomes with IFN as described above may be due to inherent differences in the animal models, delivery route, differences in IFN subtype and/or active viral antagonism of innate immunity. Since recent studies have demonstrated type III IFN to be most effective in ameliorating influenza pathogenesis in mice, comparative studies investigating the potency of different IFN subtypes should be pursued with MERS-CoV43,44,45.
Acute lung injury (ALI) in humans is well defined by a set of clinical parameters (i.e., acute onset, diffuse bilateral infiltrates on X-ray, ratio of partial pressure of arterial oxygen to inspired oxygen < 300, no evidence of elevated pulmonary arterial pressure, etc.), which can be measured in mice but require specialized procedures, equipment, and training not readily available to most researchers26,46. Moreover, animal models of ALI typically fail to recapitulate all pathologic features observed in humans likely due to differences in underlying anatomy, physiology, immunology, genetics, and complex comorbidities
联系人:李经理
手机:13621067991(wechat)
电话:13552979007(wechat)
邮箱:chemsynlab@163.com, zhangchao@chemsynlab.com
地址: 北京市大兴区金苑路26号1幢4层411室
 chemsynlab
chemsynlab